Нервная система и органы чувств
04.07.2010 13:44 | Автор: Editor |
  
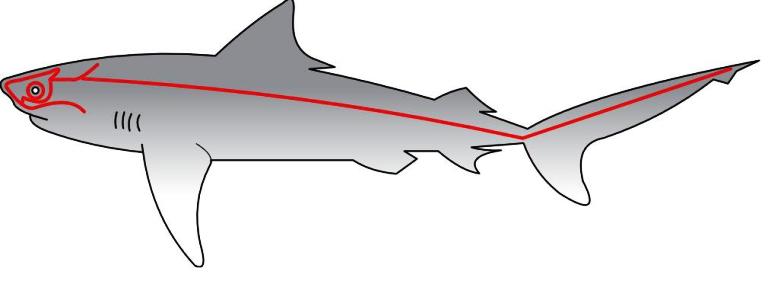
Спинной мозг проходит внутри нервных дуг позвонков по всей длине позвоночника рыбы. Аналогично миомерам и позвоночнику, в строении спинного мозу наблюдается сегментация. В каждом сегменте тела сенсорные нейроны входят в спинной мозг через дорсальные корешки, а двигательные нейроны выходят из него через вентральные. Глаза рыб по своему строению очень похожи с глазами других позвоночных. Главное принципиальное отличие рыбьего глазу заключается в том, что для фокусирования на предмете, рыбы не изменяют кривизну хрусталика, а приближают или отдаляют его от роговицы. Структура сетчатки варьирует для рыб в зависимости от места их обитания: у глубоководных видов глаза приспособлены для восприятия света преимущественно красной части спектра, а рыбы, которые живут на мелководье, воспринимают более широкий спектр. Нюх и вкус позволяют рыбам ориентироваться в химическом составе окружающей среды. Способность рыб к ощущению химических сигналов хорошо иллюстрируется лососями, которые идя на нерест из моря к речным системам, определяют по вкусу воды именно тот ручей, в которой когда-то самые вышли из икры. Обонятельные рецепторы рыб расположены в ноздрях (которые, в отличие от других позвоночных, не соединяются с носоглоткой), а вкуса — не только в ротовой пустоте, но часто и на жаберных структурах, усиках, и даже на плавниках и на коже. Главными механорецепторами рыб являются органы слуха, которые функционируют как орган и слуха, и равновесия, а также органы боковой линии. Внутреннее ухо пластинчастожабернsх (акул и скатов) и костистых рыб состоит из трех полукруглых каналов, расположенных в трех взаимно-перпендикулярных плоскостях, и трех камер, каждая из которых вмещает отолиты. Некоторые виды рыб (например, серебряный карась и разные виды сомов) имеют комплекс косточек, которые называются Вебберов аппарат и соединяют ухо с плавательным пузырем. Благодаря этой адаптации внешние вибрации усиливаются плавательным пузырем, как резонатором. Отолит в третьей камере, обеспечивает рыбе ориентацию в пространстве. Перемещение воды по поверхности рыбы ощущается структурами, которые называются нейромасты. Эти органы могут быть рассеяны поодиночке, или собраны под чещуей в совокупности каналов, которые называются боковой линией. Нейромасты включают гелевой консистенции полушарие (капулу) и сенсорные волосовидные клетки, а также синапсы нервных волокон, которые образовываются на волосовидних клетках. Перемещение воды служит причиной отгиба волосовидных клеток, который превращается в нервные импульсы. Эти импульсы позволяют составить довольно подробную картину окружающей среды: некоторые виды рыб, лишенные глаз, целиком ориентируются и перемещаются, полагаясь только на органы боковой линии.
Обновлено (10.12.2010 22:49) Пищеварительная система и питание
04.07.2010 14:10 | Автор: Editor |
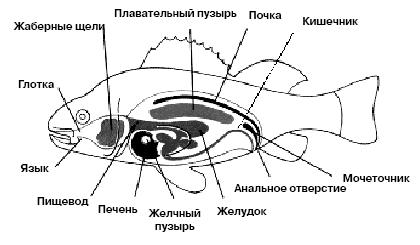
Рыбам присущ широкий спектр пищевых объектов и методов питания. В целом все рыбы могут быть разделены на травоядных, хищников, детритофагов и всеядных. Растительноядные рыбы могут питаться как макро-так и микроводорослями, а также водными цветочными растениями. Некоторые из таких рыб приспособлены к питанию планктоном, фильтруя его специализированными жаберными тычинками на жаберных дугах: например, разные виды толстолобиков (Hypophthalmichthys molitrix, Hypophthalmichthys nobilis) питаются исключительно за счет этого ресурса, и являются строго определенными рибами-фильтраторами микроскопических водорослей, которые живут в толще воды. Морские рыбы из семейства Pomacentridae питаются бентосными макроводорослями, причем каждая рыба имеет определенный участок, где она вырывает все водоросли, оставляя для роста только те виды, которыми питается — при этом не вырывая их во время питания, а лишь частично объедая. Хищные рыбы используют в качестве пищевого ресурса широчайший спектр объектов. Планктонные фильтраторы (такие как Китовая акула) отфильтровывают зоопланктон из толщи воды; кроме того, рыбы могут питаться ракообразными, моллюсками, плоскими, круглыми и кольчатыми червями, а также другими рыбами. Наиболее специализированными из хищных рыб являются те, которые питаются эктопаразитами других рыб (Labridae, Chaetodontidae и прочие), которые выбирают паразитов и отмершие кусочки кожи с поверхности тела рыб-«клиентов», которые специально посещают места обитания «чистильщиков». Много видов рыб могут изменять тип питания на протяжении жизни: питающиеся планктоном в молодом возрасте, могут переходить на питание рыбами или крупными беспозвоночными во взрослой фазе развития.
Обновлено (10.12.2010 22:49) Размножение
04.07.2010 14:14 | Автор: Editor |
Типы размножения Двуполое размножение Двуполое размножение является наиболее обычной и широко распространенной его формой. При этом способе репродукции, самки и самцы внутри вида являются четко отделенными. При этом некоторые виды могут демонстрировать очень ярко выраженные вторичные половые признаки, или половой диморфизм. Эти характеристики вторичных половых признаков обычно проявляются только одним полом (в большинстве случаев — у самцов), не относятся к половому созреванию, могут интенсифицироваться на протяжении брачного сезона, не оказывают содействие индивидуальному выживанию. Вторичные половые признаки могут проявляться в виде различий в размерах тела, частей тела (например, удлиненные плавники), строения тела (например, выросты на голове), расположении зубов, окраске, а также встречаются в виде отличий между акустическими, химическими, электрическими и характеристиками самцов и самок. Двуполый способ размножения может включать в себя моногамию, полигамию и промискуитет. Гермафродитизм Второй способ размножения рыб включает в качестве элемента изменение пола особями одного вида, когда рыбы могут функционировать то как мужская, то как женская особь случайно или последовательно. Последовательное функционирование проявляется в виде функционирования как самцов на протяжении одной части жизни, и как самок — на протяжении другой. Существуют две формы последовательного изменения пола — протоандрия и протогения. Протоандрические гермафродиты — это особи, которые в начале своей жизни являются самцами, а позднее претерпевают кардинальные перестройки половой системы и становятся полностью функциональными самками. Такая форма преобразования пола широко распространена в семействе морских окуней (Serranidae). Все губаны (Labridae) являются протогеническими гермафродитами, когда все самцы являются преобразованными с возрастом самками. В этом семействе на смену пола могут влиять как факторы окружающей среды, так и социальные отношения в популяции. Социальная структура губанов заключается в наличии гаремов, которые состоят из самок и одного большого самца. Внутри группа структурирована по размеру, с самцом на верхушке иерархии. Если изъять из группы самку, другие самки (низшие по рангу) будут изменять свое иерархичное положение, обычно сдвигаясь на одну позицию вверх. Если же изъять из группы самца, самая крупная самка гарема старается занять его место, агрессивно отгоняя самцов, которые контролируют другие гаремы. Если ей это удается, и никому из окружающих самцов не удается присоединить этот гарем к собственному, то эта самка начинает демонстрировать поведение самца, и после около 14 дней ее половая система полностью изменяется, начиная продуцировать мужские половые клетки. В таксонах, где половая принадлежность обусловлена социальной структурой, процесс изменения пола широко варьирует, и одна и та же особь может изменять пол несколько раз на протяжении жизни. С другой стороны, существуют таксоны (например, полосатые окуни, желтый окунь, большинство групперов) где половая принадлежность особей чередуется, но не испытает влияние социальной структуры. Случайные гермафродиты могут продуцировать как яйцеклетки, так и сперматозоиды — они потенциально имеют возможность самооплодотворения. Известные лишь три вида из отряда Cyprinodontiformes, которые функционируют как самооплодотворяющие гермафродиты: два вида рода Cynolebias и вид Rivulus marmoratus. При этом самооплодотворение у Rivulus marmoratus является внутренним, и в результате приводит к появлению гомозиготных, генетически идентичных потомков. Более обычная форма случайного гермафродитизма наблюдается в родах Hypoplectrus и Serranus семейства Окуневых (Percidae). Хотя эти рыбы способны продуцировать сперматозоиды и яйцеклетки одновременно, на протяжении одного нереста они функционируют как представители только одного пола. Учитывая то, что один акт нереста может длиться несколько часов, рыбы одной пары могут обмениваться половыми ролями, и продуцировать поочередно яйцеклетки (икру) или сперматозоиды (молоки). Партеногенез Несмотря на редкость этого типа размножения среди позвоночных, несколько видов рыб прибегают к нему. По определению, партеногенез заключается в развитии яйца без оплодотворения сперматозоидом этого же вида. У рыб существует вариант этого типа размножения, при котором необходимым является общий нерест с самцами того же самого вида или других видов. При этом роль самцов заключается в продуцировании сперматозоидов, которые контактируют с икринками, но не в состоянии проникнуть через их внешнюю мембрану (хорион). Контакт со сперматозоидами выполняет роль стимула, который побуждает яйцо начать развитие. При этом сперматозоиды не вносят в яйцо своего генетического материала, то есть все потомки при таком размножении будут самками, генетически идентичными с материнской особью. Классическим примером такого гермафродитизма есть гольяны (род Poeciliopsis) и европейский серебряный карась Carassius gibelio.
Овипороз состоит в высвобождении как мужских, так и женских гамет в воду, где происходит оплодотворение. Оплодотворение может быть при этом внутренним (как у скорпен (Scorpenidae) и американских сомов Auchenipteridae), но самка после этого выбрасывает оплодотворенную икру в окружающую среду. На протяжении развития зародыш использует как желток яйца, так и имеющиеся в яйце капли жиров. При ововипорозе яйца все время находятся в организме самки, и оплодотворение является внутренним. Несмотря на нахождение яиц в теле самки, между материнским организмом не возникает плацентарного контакта или контакта кругов кровообращения. Вместе с тем зародыш во время своего развития питается запасами питательных веществ, которые имеющийся в яйце. После достижения выводкового развития, мальки выходят из яиц внутри материнского организма, после чего немедленно выходят наружу. Наиболее известными рыбами, которым присущая такая форма размножения, являются представители Poeciliidae — широко распространенные в аквариумах гуппи и меченосцы. Также данная форма размножения характерная для латемерии. Размножение в форме вивипорозу во многих чертах подобно ововипорозу, но при этом между яйцом и материнским организмом возникает плацентраний контакт или контак между их кругами кровообращения. Таким образом, эмбрион при развитии получает необходимые питательные вещества из организма матери. Наиболее обычна данная форма размножения у акул, но эта форма размножения присущая также и костистым рыбам — жителям горных водоемов из семейства Goodeidae и представителям семейства Embiotocidae. Нехарактерная для рыб забота о потомстве наблюдается преимущественно у видов в приливно-отливной зоне, в узких заливах и бухтах, а также в реках и озерах. Самец пятнадцатииглой морской колюшки сооружает своеобразное гнездо из кусочков водорослей, скрепляя их специальным секретом. Потом он загоняет в гнездо самок, которые откладывают икру, а самец остается охранять икру молодь. Самка горчака, может вытягивать своеобразный яйцеклад в длинную трубку, и откладывает икру в мантийную полость пресноводных двустворчатых моллюсков. Самец выпускает сперму рядом с моллюском, и, когда тот захватывает корм, втягивая в себя воду, и икра оплодотворяется. Особое приспособление имеется у самца австралийского гулливерова куртуса. Самец вынашивает икринки у себя на лбу. Грозди икринок, прикрепленных клейкими нитевидными выростами с обоих концов, крепко удерживаются крючком, образованным лучами его спинного плавника. Сложное поведение, связанное с заботой о потомстве, развилось у морских игл и морских коньков. Обновлено (10.12.2010 22:49) |
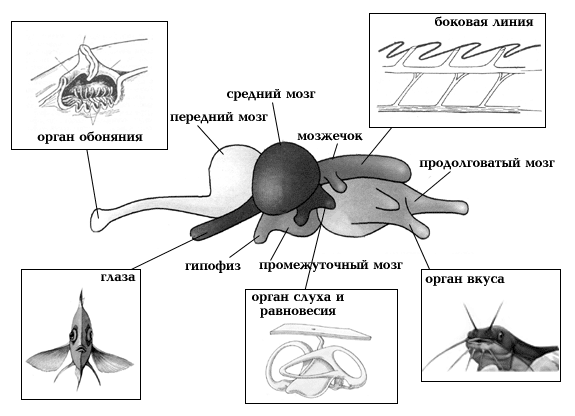
 Головной мозг рыб принято разделять на три большие части: передний, средний и задний мозг. Передний мозг состоит из теленцефалону (конечного мозга) и диэнцефалону (промежуточного мозга). На ростральном (переднем) конце теленцефалону расположенные обонятельные луковицы, которые получают сигналы от обонятельных рецепторов. Обонятельные луковицы обычно увеличены у рыб, которые активно используют нюх, например, у акул.Перемещение воды по поверхности рыбы ощущается структурами, которые называются нейромасты. Эти органы могут быть рассеяны поодиночке, или собраны под чещуей в совокупности каналов, которые называются боковой линией. Нейромасты включают гелевой консистенции полушарие (капулу) и сенсорные волосовидные клетки, а также синапсы нервных волокон, которые образовываются на волосовидних клетках. Перемещение воды служит причиной отгиба волосовидных клеток, который превращается в нервные импульсы. Эти импульсы позволяют составить довольно подробную картину окружающей среды: некоторые виды рыб, лишенные глаз, целиком ориентируются и перемещаются, полагаясь только на органы боковой линии.
Головной мозг рыб принято разделять на три большие части: передний, средний и задний мозг. Передний мозг состоит из теленцефалону (конечного мозга) и диэнцефалону (промежуточного мозга). На ростральном (переднем) конце теленцефалону расположенные обонятельные луковицы, которые получают сигналы от обонятельных рецепторов. Обонятельные луковицы обычно увеличены у рыб, которые активно используют нюх, например, у акул.Перемещение воды по поверхности рыбы ощущается структурами, которые называются нейромасты. Эти органы могут быть рассеяны поодиночке, или собраны под чещуей в совокупности каналов, которые называются боковой линией. Нейромасты включают гелевой консистенции полушарие (капулу) и сенсорные волосовидные клетки, а также синапсы нервных волокон, которые образовываются на волосовидних клетках. Перемещение воды служит причиной отгиба волосовидных клеток, который превращается в нервные импульсы. Эти импульсы позволяют составить довольно подробную картину окружающей среды: некоторые виды рыб, лишенные глаз, целиком ориентируются и перемещаются, полагаясь только на органы боковой линии.